Il y a quelques mois, j’ai écrit un court article de blog pour un projet scientifique qui expliquait en quelques paragraphes pourquoi il était à la fois si difficile et si important de modéliser les changements d’occupation des sols. En effet, on parle énormément du changement climatique alors que c’est bien la destruction des habitats naturels, et donc les changements d’occupation des sols, qui représente la menace principale pour la biodiversité à l’échelle mondiale. Je me suis dit qu’il serait intéressant de reprendre ce court article pour vous le traduire ici et l’agrémenter de quelques explications supplémentaires.
Il est aujourd’hui largement admis que le climat va fortement changer au cours des prochaines décennies, ce qui entraînera des impacts profonds sur la biodiversité et la répartition des espèces sur toute la planète. Mais, si le climat est un déterminant majeur des changements à venir, il n’est pas le seul mécanisme à la base de la distribution des espèces, ni l’unique cause de la disparition de la biodiversité. En effet, il est globalement admis que les changements d’occupation des sols représentent la première menace pour la biodiversité et sont particulièrement importants pour comprendre la distribution des espèces et des services et fonctions écosystémiques. Mais qu’est-ce qu’une « occupation du sol » et qu’est-ce que l’on entend par « changement d’occupation des sols » ?
L’usage des sols, aussi appelé occupation ou utilisation des sols (il y a quelques subtiles différences mais ce n’est pas très important ici) représente la fonction ou l’habitat qui couvre une zone définie. Par exemple, les montagnes près de chez moi sont couvertes de forêt, c’est l’habitat qui occupe cet espace. Maintenant, on peut découper plus finement les forêts en cartographiant la distribution des forêts décidues, c’est-à-dire composées d’arbres qui perdent leurs feuilles, des forêts de conifères, des forêts humides ou encore des parcelles de sylviculture etc. Les prairies naturelles ou humides, les différents types d’agriculture, les marais ou les zones urbaines et artificielles représentent aussi une occupation des sols. Chaque mètre carré pourrait être classifié dans une catégorie ou une autre d’occupation des sols.
Mais alors, de quoi parle-t-on exactement lorsque l’on évoque les changements d’occupation des sols ? C’est en fait un terme générique qui correspond généralement à la destruction d’habitats naturels dans le but d’en faire une zone plus productive ou artificielle, généralement moins diversifiée et moins habitable pour la biodiversité : on va raser la forêt pour en faire un champ de culture intensive, ou bien on va détruire un champs agricole pour en faire un supermarché. Mais cela fonctionne aussi avec une intensification des usages. Par exemple, une prairie de fauche ou un pâturage, qui abritent une diversité élevée alors même qu’elles sont directement entretenues par les humains, peuvent être transformés en cultures intensives. Les changements d’occupation des sols pourraient aussi fonctionner dans l’autre sens : un parking de supermarché est renaturé et transformé en forêt, mais ce genre de changement est tellement rare qu’il n’a absolument aucun impact capable de contrebalancer la destruction des habitats naturels. En revanche, la déprise agricole de certains territoires, ou l’abandon des pâturages peut permettre à la forêt de repousser. De manière général, l’occupation des sols change avec le temps en fonction des priorités du territoire.
Les pâturages alpins ont tendance à être abandonné ce qui permet à la forêt de pousser plus haut en altitude.
Pourquoi modéliser les changements d’occupation des sols ?
La distribution des usages des sols et des habitats est particulièrement importante pour modéliser, par exemple, la distribution des espèces. Nous avions déjà parlé dans cet article et dans celui-ci. En effet, il est pratique de savoir où se trouvent les forêts pour modéliser la distribution des espèces forestières. Il est donc très important de pouvoir cartographier un territoire pour savoir de quoi il est composé. Si vous voulez regarder un peu les cartographies numériques qui existent déjà, vous pouvez vous rendre sur le site de CORINE-land-cover ou de Google Earth Engine.
Si vous avez suivi les derniers articles publiés, vous savez d’ores et déjà que pour modéliser la distribution des espèces dans le futur, il faut avoir accès à deux choses : 1) au climat futur, en se basant par exemple sur différents scénarios de changement climatique et 2) à la distribution future de l’occupation des sols. Nous en avions longuement parlé dans cet article notamment. Mais il y a un problème, comment connaître l’occupation future des sols ? Pour le climat, les méthodologies sont relativement connues, mais pas pour l’occupation des sols. D’ailleurs, aujourd’hui encore, la majorité des articles scientifiques qui tentent de modéliser la distribution future des espèces ne se basent que sur le climat, ce qui est gravement réducteur !
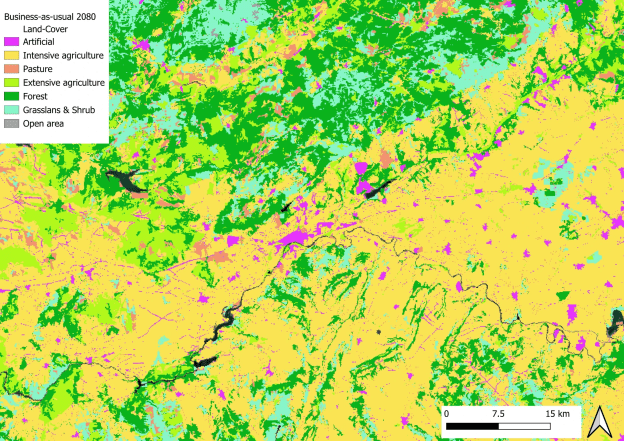
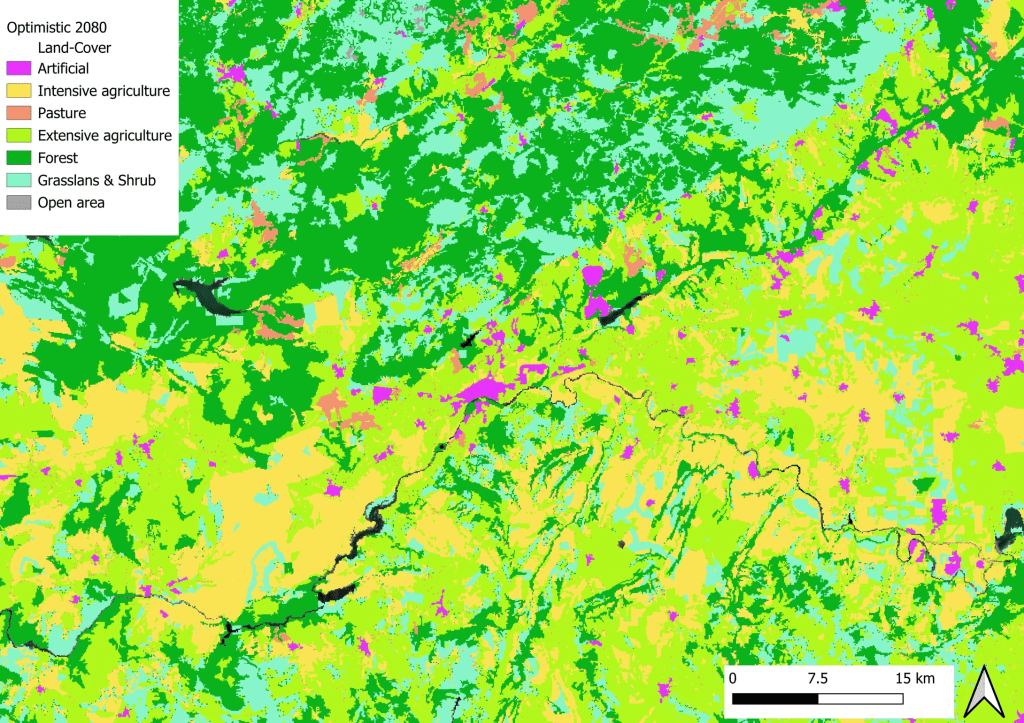
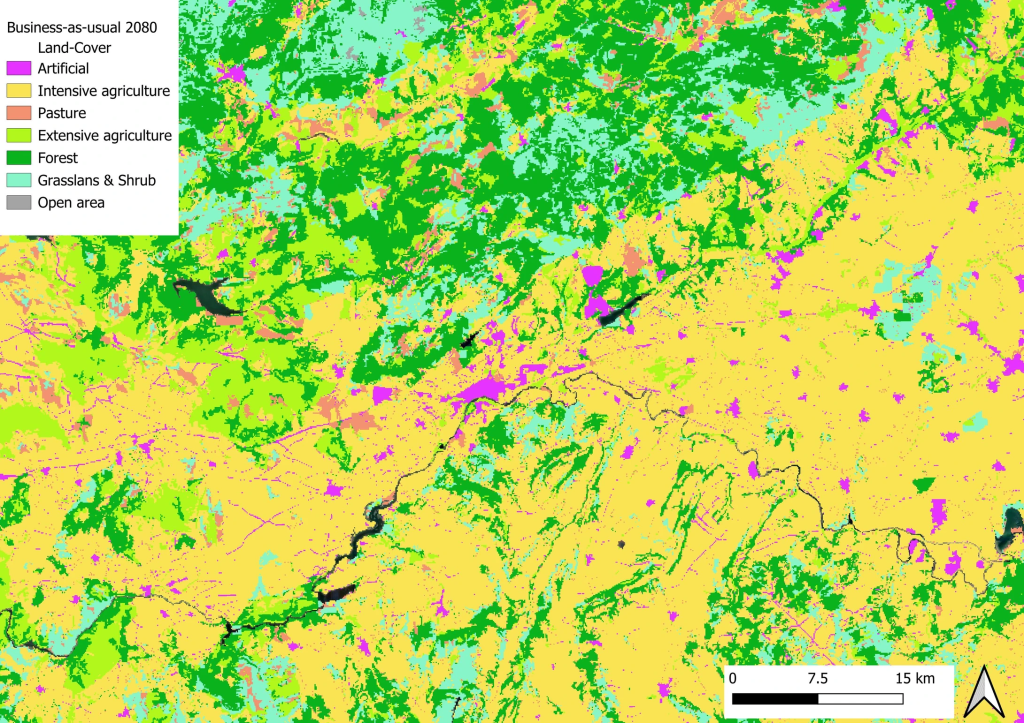
Comme nous ne pouvons pas savoir a priori comment les paysages vont changer dans le futur et s’il y aura plus ou moins de chaque catégorie d’habitat, il faut là encore jouer avec des scénarios. Par exemple, dans mon projet de recherche actuel, nous nous sommes basés sur 3 horizons plausibles définis par les acteurs de la conservation de la nature et de l’environnement. Nous avons un premier scénario extrêmement pessimiste où les habitats naturels régressent pour laisser place à des usages productifs, et où les zones extensives sont transformées en agriculture intensive. Nous avons ensuite un scénario intermédiaire qui serait une continuation de ce que l’on observe ces dernières années : une intensification des usages mais une protection de la nature existante et efficace. Enfin nous avons un scénario optimiste où les pratiques intensives ont tendance à laisser place aux pratiques plus extensives, et ces dernières sont converties en des zones plus naturelles.
Méthodologie
Bien, nous avons donc nos scénarios futurs et la cartographie de l’occupation actuelle des sols, que manque-t-il ?
Si nous souhaitons construire un modèle qui arrive à prédire les changements d’occupation des sols dans les prochaines décennies, il nous manque deux données cruciales. La première est une carte de l’occupation des sols passée, avec les mêmes catégories que la carte actuelle, afin d’étudier les tendances et les changements qui ont eu lieu ces dernières décennies. La seconde est une collection de données environnementales, appelées « prédicteurs », qui vont nous permettre de comprendre et de projeter les changements dans le futur. Je m’explique.
En comparant une carte ancienne avec sa version actuelle, il est possible de mesurer les changements d’occupation des sols : par exemple, en 20 ans, les aires urbaines ont augmenté de 10% alors que les prairies naturels ont diminué de 30%. Cela est particulièrement intéressant pour projeter ces changements dans le future : dans 20 ans, on peut imaginer qu’il y aura aussi +10% d’aires urbaines et -30% de prairies naturels, si l’on suit un scénario de continuation. Mais dans notre projet, cela n’est pas vraiment pertinent car nous avons développé des scénarios qui nous donnent des directives qui peuvent être très différentes de ce qui a été observé par le passé. Et puis rien ne nous dit que les changements futurs seront dans la continuité de ceux qui ont eu lieu par le passé… Non, ce qui nous importe vraiment dans la comparaison de ces deux cartes, ce sont les zones où les changements ont eu lieu et leurs caractéristiques. C’est cela qui nous souhaitons comprendre et modéliser.
En effet, en utilisant les prédicteurs environnementaux tel que la proximité avec des infrastructures de communication comme des routes, ou des villes, la pente, l’altitude, la distance aux zones sauvages et tout un tas d’autres paramètres, on peut alors prédire les zones qui ont le plus de chance de changer dans le futur. Prenons deux exemples. En analysant où les zones urbaines ont été construites par le passé, on peut alors conclure que les nouveaux bâtiments et nouvelles infrastructures sont généralement construits dans des zones plutôt plates, proche des routes et d’autres centres urbains, mais plutôt loin des forêts ou des parcs naturels. Cela nous donne des indications sur les possibles zones de transition vers de nouvelles aires urbaines dans le futur. Ainsi, une zone qui comporte toutes ces caractéristiques a de grandes chances de se transformer en milieu urbain dans les années à venir, si nous considérons dans notre scénario que les zones urbaines vont effectivement s’étendre dans le futur. De même, nous pouvons voir que les nouvelles forêts ont plutôt poussé proche des zones déjà boisées, naturelles ou agricoles, et plutôt loin des centres urbains par le passé, ce qui nous donne des indications importantes sur les potentielles nouvelles zones qui seront reboisées dans le futur.
En analysant ainsi toutes les transitions passées d’une catégorie d’occupation du sol avec une autre, on peut reconstruire une carte de probabilité de chacune de ces transitions et projeter dans le futur les changements attendus en fonction des directives que l’on donne au modèle (plus de forêt, moins d’agriculture etc.). Magique non ?
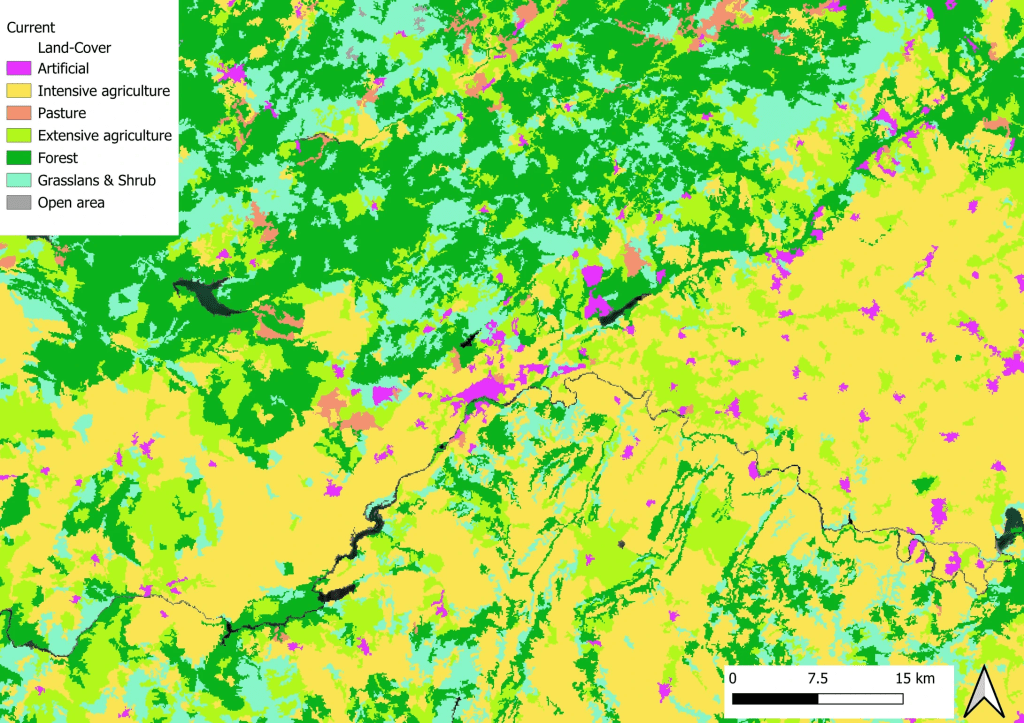
Occupation actuelle des sols près d’une petite ville du centre de l’EspagneLa même zone projetée dans 50 ans dans un futur optimiste : vous remarquez le remplacement des zones agricoles intensives (en jaune) par des zones plus extensives (en vert clair)La même zone projetée dans 50 ans dans un futur pessimiste cette fois : vous voyez ici le remplacement des zones boisées (en vert foncé) ou des prairies naturelles (en bleu clair) par des zones d’agriculture intensives (en jaune) ainsi que la multiplication des zones urbaines (en rose)
Importance des résultats et nouveaux problèmes
Maintenant que nous avons nos cartes futures d’occupation des sols, à quoi peuvent-elles bien nous servir ? Et bien comme expliqué précédemment, ces cartes peuvent servir de base pour étudier la distribution future des espèces, et voir comment les changements climatique et d’occupation des sols affectent les plantes et animaux de nos territoires. C’est exactement ce que j’ai fait dans mon 3e chapitre de thèse que j’ai résumé ici. On peut alors prédire la vulnérabilité des espèces, voir quelles sont les plus menacées par ces changements environnementaux, et prendre des mesures de conservation avant qu’il ne soit trop tard. On peut faire la même chose avec la distribution future des services écosystémiques ou la connectivité du territoire afin de mesurer précisément ce que l’on gagne et ce que l’on perd en fonction de la vision que nous avons d’un territoire. Grâce à ces données, on peut par exemple tester l’efficacité des zones protégées dans le futur ! Bref c’est un outil fantastique qui nous permet d’être plus précis dans nos résultats mais qui est aujourd’hui trop peu utilisé par les écologues… Peut-être à raison.
En effet, projeter le climat dans le futur grâce à des modèles est très complexe et engendre tout un tas d’approximations, certes. Mais ces projections se basent sur de la physique du climat, c’est palpable et mathématique. Les projections d’occupation des sols ne se basent que sur des scénarios imaginaires que nous avons inventé alors que la réalité est très politique. Il est ainsi virtuellement impossible de modéliser avec certitude l’organisation des paysages de demain, alors que c’est possible pour le climat en considérant quelques hypothèses de base. Les changements d’occupation des sols dépendent de décisions politiques et géopolitiques, comme l’approvisionnement en matières premières, le soucis de la couleur politique au pouvoir concernant l’écologie, la résilience, la souveraineté alimentaire et d’autres événements impossibles à prévoir. Pour se débarrasser de ce problème, on utilise généralement des scénarios très différents pour imaginer tous les futurs possibles à l’échelle d’un territoire, mais le détail reste de toutes manières impossible à prévoir. Par exemple, on peut imaginer sans trop d’incertitudes que plusieurs hectares de forêt seront rasées dans les prochaines années, mais prédire exactement où au mètre près est impossible. Pour tenter une nouvelle fois de prendre en compte ce soucis du détail, une technique consiste à faire plusieurs réplicats, des dizaines, voire des centaines, pour chaque scénario, avec des changements modélisés à différents endroits du territoire histoire de prendre en compte notre incapacité à produire une carte réelle de l’occupation futur des sols. Mais les résultats de centaines de réplicats sont difficiles à résumer sans perdre de l’information et encore plus difficiles à communiquer.
Bon, ces différentes techniques permettent au moins d’analyser les grandes tendances et d’en tirer des conclusions générales qui peuvent aider à prendre de meilleures décisions quant à l’avenir d’un territoire afin de mieux conserver la biodiversité dans le futur, de sauvegarder certains services écosystémiques importants ou de planifier une restructuration agricole. Mais cet exercice reste majoritairement académique et possède des incertitudes difficiles à traduire dans le monde réel.
J’espère que ce nouvel article vous a plu ! J’essaie de reprendre le temps de rédiger des petits billets sur des sujets finalement peu ou pas du tout traités dans les médias classiques de vulgarisation scientifique/écologique afin d’aider de potentiels étudiants ou curieux à mieux comprendre le travail d’écologue, n’hésitez pas à me dire si vous souhaitez que je traite un sujet en particulier ou à me contacter si vous souhaitez plus d’information !
D’autres articles sont en court de rédaction et devraient sortir dans les prochains mois, restez connectés !
Nous savons maintenant de manière certaine que les changements climatique d’une part et d’utilisation des sols d’autre part vont avoir un impact important sur la distribution des espèces dans les décennies à venir. Cela paraît logique : si une plante s’est adaptée à certaines conditions environnementales, comme le climat ou le type d’habitat, et que ces conditions changent ou disparaissent, elle a moins de chance de survie sur le long terme. Mais comment mesurer la vulnérabilité des espèces aux changements futurs ? Comment vont évoluer les distributions des espèces ? Quelles sont les plus vulnérables et qu’ont-elles en commun ? Existe-t-il des espèces qui, au contraire, vont bénéficier des changements environnementaux ? En étudiant les changements attendus dans la distribution d’environ 1700 espèces de plantes de tous types, nous avons pu répondre à ces questions, et les résultats sont pour le moins surprenants…
La première partie traite de l’infrastructure écologique, le cadre conceptuel de mon doctorat, et est disponible en cliquant ici
La seconde partie explique comment modéliser la distribution des espèces et est plutôt importante pour bien comprendre la méthodologie utilisée dans ce chapitre. Elle est disponible en cliquant ici
Pour ce troisième volet, je vais me baser sur un article publié et disponible ici : https://onlinelibrary.wiley.com/doi/full/10.1111/jbi.15132 . Cet article est le cœur de mon travail doctoral, il est donc très dense et très complexe. Je vous présente ici un résumé vulgarisé sans entrer dans les détails méthodologiques qui n’ont pas vraiment d’importance. En revanche, pour bien comprendre le contexte et la méthode derrière les modélisations de distribution d’espèces, je vous conseille de lire (ou d’écouter!) les deux premières parties publiées sur le site. Vous pouvez aussi creuser certaines parties ou accéder aux sources dans l’article scientifique qui a été publié.
Modéliser la distribution d’espèces dans les conditions présentes est déjà assez compliqué et requiert beaucoup de prudence et de compréhension de l’écologie des espèces d’intérêt. Mais quand il s’agit d’appliquer cette méthode à des conditions qui n’existent pas ou pas encore, comme c’est le cas avec le changement climatique, c’est en fait tout bonnement impossible… À moins d’utiliser une petite technique. En effet, nous en avons déjà longuement parlé dans la partie 2 du résumé de ma thèse, les modèles de distribution essaient de calculer la niche écologique réalisée des espèces en se basant sur des variables environnementales et des points d’observation géoréférencés. Sauf que dans notre cas, nous ne savons pas où se trouveront les espèces dans le futur, et nous ne savons pas non plus quel sera exactement le climat de notre aire d’étude. Pour répondre à ces deux problématiques majeures il faut effectuer deux ajustements :
Travailler avec des scénarios. Comme nous ne connaissons pas le climat futur, il faut viser large et utiliser plusieurs types de modélisations climatiques et plusieurs scénarios. Nous avions déjà parlé de ces scénarios dans cet article. En effet, certains scénarios dits « optimistes », partent du principe que nous allons collectivement drastiquement réduire nos émissions à partir de… il y a quelques années. Aïe…. Bon, ce n’est pas très réaliste mais peu importe, l’idée est avant tout de voir les conséquences d’un scénario extrêmement optimiste où nous redoublons d’effort pour rattraper notre retard (même si vous avez compris, je ne crois pas du tout que nous y arriverons). Il existe aussi des scénarios « pessimistes », dont nous empruntons la voie jusqu’à preuve du contraire, où la concentration en CO2 continue d’augmenter jusqu’à la fin du siècle. Le fait d’utiliser ces deux scénarios « extrêmes » nous donne une idée de ce qu’il devrait se passer, c’est-à-dire quelque chose quelque part entre les deux.
Étudier la niche écologique complète des espèces. La petite astuce est de déterminer où se trouvent aujourd’hui les régions qui ont un climat similaire à celui de notre zone d’étude (ici Genève et ses alentours) dans le futur. Généralement, sous nos latitudes, il faut regarder du côté du sud de la France, de l’Espagne ou de l’Italie puisque le climat attendu à Genève en 2050 sera similaire à des villes de ces pays. Ensuite, il faut voir si les espèces qui poussent actuellement à Genève sont aussi capables de pousser dans ces régions plus chaudes. En effet, une plante que l’on trouverait de la France jusqu’au Danemark sera bien moins adaptée aux conditions futures plus chaudes qu’une autre que l’on trouverait entre l’Espagne et la Suisse. Cela paraît logique, mais pour le prendre en compte, il faut étudier la distribution des espèces à une échelle spatiale bien plus grande. Dans ce cas, nous avons pris en compte tous les climats d’Europe.
En résumé, nous avons collecté plus de 22 millions d’observations précises de plantes à l’échelle de toute l’Europe pour environ 1700 espèces présentes dans le Grand Genève (territoire transfrontalier entre la France et la Suisse), et nous avons modélisé leur distribution pour la situation présente, ainsi qu’en 2050 dans des scénarios optimistes de changement climatique et dans des scénarios pessimistes. Pour les distributions futures, nous avons aussi pris en compte les changements d’utilisation des sols qui ont été modélisés exprès pour cet exercice (je vous reparlerai prochainement de cette méthode et de son utilité).
Au total et pour chaque espèce, nous avons donc la distribution actuelle ainsi que les distributions futures dans deux scénarios différents. Nous avons aussi accès à d’autres informations comme par exemple le niveau de menace de chacune des espèces ainsi que leur origine (indigène, c’est-à-dire locale, ou exotique, ce qui signifie introduite). Il est donc possible, grâce à l’ensemble de ces informations, de mesurer précisément comment les distributions vont changer entre le présent et le futur en fonction des changements globaux (climatique et d’utilisation des sols), de voir la différence entre les scénarios les plus optimistes et les plus pessimistes, et de comparer ces résultats en fonction du statut de menace ou de l’origine des espèces. Grâce à ce travail, nous pourrons alors répondre aux questions suivantes :
Est-ce que les espèces aujourd’hui menacées d’extinction sont plus vulnérables aux changement globaux que les espèces dont les populations sont en bonne santé ?
Existe-t-il une différence de vulnérabilité entre les espèces indigènes et introduites ?
Quelle menace aura le plus d’impact sur la vulnérabilité des espèces, le changement climatique ou d’utilisation des sols ?
Quel est l’impact direct de notre (in)action climatique sur la possibilité de survie des espèces dans les prochaines décennies ?
Jolie promesse non ?
Méthodologie utilisée dans cet article pour la modélisation des espèces
Méthode
Je vous passe la méthodologie utilisée pour modéliser la distribution des espèces, elle est résumée dans la figure ci-dessus et plus largement développée dans la partie 2 du résumé de ma thèse. C’est assez incompréhensible mais n’hésitez pas à me contacter si vous souhaitez des détails. Partons donc du principe que nous avons déjà les résultats, c’est-à-dire les distributions actuelle et futures dans deux scénarios de changement climatique : optimiste et pessimiste. Nous avons alors créé 6 indicateurs pour mesurer comment les changements globaux vont impacter la distribution de nos 1700 espèces de plante.
La taille de la distribution. Si la distribution future est plus grande que la présente, alors l’espèce sort gagnante des changements globaux. Au contraire, si elle est plus petite, cela signifie qu’elle en train de se raréfier ou de disparaître.
La qualité du milieu. Il n’y a pas que la taille qui compte, la qualité est tout aussi primordiale :)). Ainsi, une espèce peut avoir une distribution identique dans le futur, mais les changements de conditions environnementales font que la zone est moins propice à sa croissance. Par conséquent, la compétitivité de l’espèce va baisser et la plante peut se raréfier.
La répartition de l’espèce. Cela représente la distribution relative de l’espèce par rapport à la surface totale de l’aire d’étude. Cet indice est très important car une espèce très rare sur le territoire est d’autant plus vulnérable aux changements de conditions et plus à même de disparaître localement qu’une espèce à la répartition très large.
La fragmentation. On mesure ici l’évolution de la fragmentation de la distribution des espèces. Si la distribution est plus fragmentée, c’est-à-dire découpée par des zones non habitables, alors l’espèce est plus menacée par les changements de conditions que si elle avait une distribution continue sans obstacle.
Indice de migration. Si la distribution future se superpose parfaitement avec celle d’aujourd’hui alors l’espèce est moins vulnérable que si elles ne se superposent pas. Dans ce cas, l’espèce doit migrer et s’installer dans de nouvelles zones, ce qui n’est pas toujours facile dans un lapse de temps aussi court. Une migration « forcée » importante est donc globalement négatif pour la pérennité de l’espèce. Au contraire, si les zones actuellement favorables au développement de l’espèce le restent dans le futur, elle n’est pas vulnérable au changement de conditions.
Le potentiel de colonisation. Si l’espèce est en capacité de coloniser de nouvelles zones dans le futur, cela est plutôt bénéfique pour sa survie et réduit sa vulnérabilité aux changements environnementaux.
Une fois l’ensemble de ces indicateurs calculés, il nous faut pouvoir les résumer pour qu’ils soient plus faciles à comprendre. On crée alors un indice de vulnérabilité globale qui va de 0 pour les espèces qui vont énormément bénéficier des changements globaux (distribution agrandie, qualité du milieu améliorée, habitats mieux connectés etc.), à 9 pour les espèces extrêmement vulnérables (distribution plus petite, qualité moindre, fragmentation accrue etc.). Vous trouverez la méthode exacte de calcul de cet indice dans l’article scientifique. Les résultats sont quelques peu surprenants, essayons de les décortiquer et de les discuter.
Résultats
Distributions actuelle et futures
Les résultats montrent qu’à l’horizon de 2050, 4 espèces de plantes auront disparu de la région genevoise. C’est à la fois peu comparé à notre jeu de données et en même temps, en moins de 3 décennies, c’est énorme. Nous avons aussi remarqué que certaines espèces devraient énormément profiter des changements de conditions comme par exemple l’orchidée méditerranéenne Himantoglossum robertianum ainsi que Gleditsia triacanthos. Ces deux espèces sont bien différentes mais illustrent parfaitement les espèces gagnantes du changement climatique. La première est une plante qui vient des climats chauds méditerranéens, il est donc normal qu’avec la hausse des températures, elle ait davantage de zones habitables à Genève et aux alentours. Elle est d’ailleurs naturellement en train de coloniser la France en remontant vers le Nord, maintenant que l’hiver n’est plus aussi rude qu’avant (s’il fallait une nouvelle preuve du réchauffement climatique). La seconde espèce est un arbre exotique, c’est-à-dire qu’il vient d’une autre région que la nôtre. Dans ce cas, il est natif du sud des Etats-Unis et du nord du Mexique où le climat est plus chaud et plus sec que chez nous. Il est donc parfaitement normal qu’il y trouve son compte avec les changements de conditions.
Les distributions futures seront différentes des actuelles pour la majorité des plantes. Voici quelques résultats en vrac lorsque l’on regarde les valeurs moyennes de notre jeu de données :
Pas ou peu de différence d’habitabilité entre le présent et le futur, cela signifie que les zones habitables futures seront globalement de qualité similaire à celles habitables aujourd’hui.
Alors que les espèces indigènes ont aujourd’hui une répartition plus grande dans le territoire, la tendance montre qu’un inversement est en cours : la répartition des plantes indigènes baissent dans le futur alors que c’est l’inverse qui est observé pour les exotiques qui gagnent donc du terrain.
En lien avec le résultat précédent, l’aire de distribution des espèces locales va globalement baisser, l’intensité étant très variable en fonction des espèces, alors que celle des espèces exotiques va fortement augmenter, d’environ 50 km carré en moyenne pour les scénarios optimistes et d’environ 75 km carré pour les scénarios pessimistes !
Les espèces indigènes vont devoir migrer pour trouver des habitats favorables, surtout dans les scénarios pessimistes, alors que ce ne sera pas du tout le cas pour les espèces exotiques dont les milieux vont rester favorables à leur développement.
Le potentiel de colonisation de nouvelles aires est très élevé pour les espèces exotiques alors qu’il est faible pour les espèces locales.
Enfin, alors que les espèces indigènes vont voir leur aire de distribution se fragmenter de plus en plus dans le future, c’est tout l’inverse qui est observé pour les espèces exotiques.
Vous l’aurez compris, les indices sont globalement (très) négatifs pour les espèces locales, indigènes, alors qu’ils sont (très) positifs pour les espèces exotiques déjà présentes sur le territoire. Autre résultat intéressant, les tendances observées sont toujours accentuées dans les scénarios pessimistes de changement climatique : une espèce en déclin faible dans les scénarios futurs optimistes sera en déclin fort dans les pessimistes, mais l’inverse fonctionne également, une espèce qui bénéficie moyennement des changements globaux dans les scénarios optimistes sera beaucoup plus avantagée dans les pessimistes. On pourrait être tenté de penser que ces résultats ne sont pas si grave et que la disparition d’espèces locales sera contrebalancé par le développement des exotiques. Malheureusement, ce n’est pas si simple et l’on décortique ce sujet plus longuement dans la discussion.
Himantoglossum robertianum observé en fleur autour de Montpellier en Février 2017. Cette espèce sera de plus en plus commune dans la région de Genève et dans toute la France dans les prochaines décenniesArum italicum va aussi bénéficier des changements globaux
Indice de vulnérabilité
Si l’on regarde la valeur de l’indice de vulnérabilité pour l’ensemble des plantes de notre jeu de données, les résultats confirment une tendance inquiétante mais prévisible : en moyenne, les plantes sont plus vulnérables dans les scénarios pessimistes que dans les scénarios optimistes de changement climatique. Cela signifie que notre capacité à réduire nos émissions de gaz à effet de serre a un impact direct sur la possibilité de survie de plantes à l’échelle locale. Un autre résultat intéressant de cette étude est en réalité un non résultat : il n’y aucune différence de vulnérabilité entre les espèces déjà menacées d’extinction et les espèces dont les populations sont en bonne santé. Ce résultat a plusieurs conséquences : premièrement, les changements climatique et d’utilisation des sols impacteront toutes les espèces, qu’elles soient déjà menacées ou en bonne santé n’y changera rien; deuxièmement, les espèces aujourd’hui en danger d’extinction le sont pour d’autres raisons puisque les changements globaux ne semblent pas nécessairement aggraver la situation et enfin; les statuts de menace de l’IUCN ne nous renseignent pas sur la vulnérabilité des espèces aux changements globaux, et c’est un peu embêtant. Là aussi, on développe un peu plus ces idées dans la discussion car il y a beaucoup à dire.
En gros, les espèces indigènes sont vulnérables aux changements globaux alors que les espèces exotiques bénéficient de ces changements de conditions, et cette tendance est statistiquement plus forte dans les scénarios pessimistes de changement climatique. Pour vous donner une idée de l’ampleur de cette vulnérabilité, plus d’une espèce sur 5 est modérément vulnérable (plus de 350 espèces) dans les scénarios optimistes et cela monte à plus d’une espèce sur 3 (580 plantes) dans les scénarios pessimistes. Si l’on regarde maintenant les espèces extrêmement vulnérables, on a environ 5% (80 espèces) pour les scénarios optimistes et 12% (200 espèces) pour les scénarios pessimistes. Nulle doute qu’une partie d’entre-elle va localement disparaître dans les prochaines décennies. En revanche, nous l’avons vu, les changements de conditions n’ont pas que des effets négatifs et l’on compte aussi beaucoup d’espèces bénéficiaires : un peu moins de 19% des plantes (environ 320 espèces) montrent des bénéfices modérés dans les scénarios optimistes et environ 1 quart des espèces (440 espèces) dans les scénarios pessimistes de changement climatique. Pour les plantes qui vont fortement bénéficier de ces changements, elles représentent un peu moins de 4% des plantes étudiées dans les scénarios optimistes et moins de 7% dans les pessimistes. Nous voyons que le nombre d’espèces avantagées par les changements globaux et le nombre de celles désavantagées sont assez proches, même si nous avons un nombre significativement plus élevé d’espèces vulnérables.
Si les chiffres exacts vous intéressent, je vous invite à lire l’article scientifique dont le lien est posté en haut de cette page 🙂
Campanula scheuchzeri, absolument pas menacé d’extinction dans la région genevoise, elle est en revanche extrêmement vulnérable aux changements globauxCirsium spinosissimum, emblématique des prairies alpines, est aussi très vulnérable aux changements globauxDactylorhiza sambucina, une orchidée des prairies de moyenne altitude est aussi très vulnérable aux changements globaux
Discussion
Bon, maintenant que je vous ai noyé sous une montagne de chiffres en bon scientifique que je suis, prenons un peu de recul et résumons les résultats de cette étude avant d’analyser les tendances dans leur globalité.
Les effets des changements globaux varient, certaines espèces y gagnent, majoritairement des espèces exotiques introduites, alors que d’autres, plus nombreuses, y perdent. Les espèces qui y perdent sont majoritairement les espèces locales qui représentent l’écrasante majorité des plantes dominantes de nos forêts, prairies, et milieux humides. Aussi, les effets sont exacerbés dans les scénarios pessimistes de changement climatique que nous suivons actuellement (jusqu’à preuve du contraire, vous connaissez l’adage). Cela signifie que nos actions (et plus précisément notre inaction) pour lutter contre le changement climatique ont un réel impact sur la survie des plantes de nos régions !
Avant de discuter un peu plus de ces résultats et de ce qu’ils signifient, il est toujours bon de rappeler qu’ils ont été obtenus à partir de modèles, qui ne sont, par définition, jamais parfaits. On en parlait d’ailleurs déjà dans le chapitre précédent. Pour vous donner un exemple, on ne peut littéralement pas prédire une catastrophe naturelle qui accélérerait le phénomène (ou pas, s/o le COVID), ou bien l’arrivée au pouvoir d’un-e climatonégationniste qui décide que, finalement, l’écologie et la protection de l’environnement ben on s’en tape. Ici, on ne peut que regarder ce qu’il se passe aujourd’hui et l’extrapoler linéairement dans le futur afin d’avoir une idée de la moyenne de ce qu’il devrait se passer, quand bien même on sait que les changements se font plutôt par marches d’escalier (une énième sécheresse intense et toute une espèce disparaît d’une région d’un coup, par exemple). Aussi, il est toujours bon de rappeler qu’il est très difficile de prendre en compte la résilience des écosystèmes ou les intéractions entre espèces qui peuvent à la fois ralentir les effets des changements globaux car les habitats modifient le climat localement, ou les accélérer drastiquement à cause des liens entre les espèces d’une même chaîne trophique par exemple.
Discutons de deux résultats importants de cette étude.
Tout d’abord, nous avons vu que les espèces indigènes sont globalement vulnérables aux changements globaux alors que les espèces exotiques devraient plutôt en bénéficier. Lorsque l’on regarde les chiffres, on pourrait presque penser que finalement, comme on a grosso modo autant d’espèces vulnérables qu’avantagées, alors tout va s’équilibrer et tout devrait s’arranger. Bon déjà ce n’est pas le cas puisque dans notre jeu de données, nous trouvons une majorité d’espèces vulnérables, mais admettons. Déjà, nous ne sommes pas sûr que les espèces vulnérables et bénéficiaires se trouvent dans les mêmes habitats. Si les plantes qui disparaissent se trouvent majoritairement en prairie, mais que les espèces qui bénéficient des changements globaux se trouvent en forêt, ça ne fonctionne pas. Dans notre cas c’est d’ailleurs ce que l’on observe : les plantes les plus vulnérables vivent dans les prairies subalpines et celles qui vont bénéficier des changements globaux vivent en plaine. Ensuite, et nous en avions déjà parlé dans plusieurs articles, notamment celui-ci, les espèces comme les écosystèmes ont des fonctions qui permettent un certain équilibre du milieu. On ne peut donc pas simplement remplacer une espèce par une autre si leurs fonctions ne sont pas identiques. Prenons un exemple très réel et terriblement résumé. Remplacer un ver-de-terre local qui recycle les nutriments par un vers plat introduit et prédateur a toutes les chances de bousculer les écosystèmes. C’est exactement ce qui en train de se passer avec l’espèce exotique Obama nungara qui est prédatrice des vers-de-terre de chez nous. Deux espèces qui paraissent très similaires (en réalité elles sont plutôt éloignées génétiquement mais c’est pour l’exemple) ne peuvent pas pour autant s’inter-changer. Là où le problème est encore plus grand dans cette étude, c’est bien l’origine des espèces (clin d’œil à Darwin), puisque ce sont les exotiques qui seront avantagées dans le futur, au détriment des indigènes. Et les espèces exotiques n’ont par définition pas évolué dans le nouveau milieu où elles se trouvent et cela peut avoir de lourdes conséquences, notamment à cause de leur manque d’intéraction avec les autres espèces locales. Par exemple, elles peuvent ne pas être mangées, elles ne servent donc pas de nourriture aux animaux et s’extirpent des chaînes trophiques, elles peuvent ne pas être attaquées par les maladies locales qui ne les reconnaissent pas, ce qui leur confère un avantage énorme par rapport aux espèces indigènes, elles peuvent altérer le milieu durablement, modifiant des écosystèmes entiers qui n’étaient pas habitués à leur présence etc. Histoire de finir d’enfoncer le clou, les espèces exotiques de la région sont majoritairement des espèces sélectionnées, introduites et modifiées par les humains pour tout un tas de raisons, généralement esthétiques ou utiles, afin de les garder dans des pots en ville par exemple, mais vraiment pas pour des raisons écologiques qui leur permettraient de remplacer les arbres de nos forêts. Bref, on ne peut pas remplacer des espèces indigènes par des espèces exotiques, surtout pas en si peu de temps, et encore moins si ce sont des espèces que l’on a sélectionnées pour d’autres usages.
On pourrait parler de cette thématique durant une éternité, c’est littéralement un pan entier de recherche en écologie, en restauration des écosystèmes et en conservation, mais je soulève ici juste quelques points de réflexion. Bref, l’autre résultat qui, je pense, est très intéressant, est l’absence de lien entre la vulnérabilité des espèces et les statuts IUCN. Cela signifie que les changements environnementaux, en particulier le changement climatique puisque c’est le moteur principale de la vulnérabilité des espèces dans cette situation, vont affecter de la même manière des espèces rares en danger d’extinction, et des espèces communes dont les populations sont en pleine forme. On pourrait être tenté d’imaginer que le changement climatique pourrait alors aider certaines espèces à recouvrer un peu de force afin de sortir du danger d’extinction. Mais cela signifie surtout autre chose : les espèces aujourd’hui en danger d’extinction, le sont pour d’autres raisons que le changement climatique. Autrement dit, le changement climatique va ajouter une pression supplémentaire à des espèces déjà clairement menacées et il est assez utopique de penser qu’il va pouvoir aider certaines plantes à surpasser les autres problèmes environnementaux majeurs qui les impactent déjà, et à cause desquels elles sont justement en train de disparaître. Ce résultat indique aussi quelque chose d’un peu terrifiant : les statuts de menace que nous avons développé ne prennent pas en compte les changements climatiques. En réalité, et nous en avions déjà parlé dans cet article sur les plantes carnivores, les statuts de menace sont définis en se basant sur l’évolution passée et la situation actuelle des populations. Ils ne se basent absolument pas sur les menaces futures. Et ça c’est un problème : on ne peut pas correctement planifier la conservation des écosystèmes si on ne regarde que les situations passée et actuelle, d’autant plus si l’on considère le temps nécessaire pour étudier les taxons concernés, publier les résultats, changer les statuts de menace, trouver des financements pour mettre en place des mesures environnementales, faire passer les lois nécessaires, appliquer les mesures, les suivre dans le temps et étudier les impactes qu’elles ont eu sur la santé des populations afin de les ajuster etc. Cela prend un temps de dingue, comme dirait l’autre. Au contraire, prendre en compte dans ces statuts de menace la vulnérabilité aux conditions futures pourraient nous aider à prendre de meilleures décisions et plus rapidement !
Voilà tout ce que j’avais à vous dire sur cet article, je trouve personnellement super intéressant de faire ce genre d’étude pour mieux connaître les territoires et les vulnérabilités des espèces qui les peuplent. Ce n’est évidemment pas avec ce genre d’articles scientifiques que l’on est publié dans des revues prestigieuses mais ils produisent pourtant des informations très utiles pour les pouvoirs publiques afin de prendre de meilleures décisions de conservation !
Merci beaucoup et à la prochaine pour le dernier chapitre, lorsqu’il sera publié !
Pulsatilla alpina, dont la couleur des fleurs dépend de l’acidité du milieu, est très vulnérable aux changements globauxLa magnifique Viola calcarata est aussi très vulnérable
Le petit monde des plantes carnivores n’est peut-être pas si exclusif qu’il n’y parait et nous découvrons régulièrement de nouvelles espèces carnivores ou protocarnivores chez les plantes à fleurs. Pourtant, il y a une vingtaine d’années, des chercheurs ont émis l’hypothèse que certaines « mousses » pourraient elles aussi être carnivores. Analysons cette découverte et profitons-en pour divaguer autour de la carnivorie végétale en testant ses limites et comparant ses approches.
Figure 1. Les sphaignes (Sphagnum spp) sont des mousses de l’embranchement des Bryophyta et représentent l’élément constitutif des tourbières puis de la tourbe en se dégradant.
C’est quoi une « mousse » ?
Commençons par le commencement et par bien comprendre les plantes que l’on appelle communément « mousses ». Ces végétaux appartiennent au groupe des « Bryophytes » dont les représentants actuels sont morphologiquement et physiologiquement très proches des premiers végétaux à avoir colonisé les terres émergées il y a environ 500 millions d’années, semble-t-il avec l’aide de champignons commensalistes. Les Bryophytes n’ont pas de racines, pas de système vasculaire et ne produisent ni fleurs ni graines, elles se disséminent grâce à des spores. Parmi les Bryophytes, il existe notamment l’embranchement des Bryophyta (les « vraies » mousses) dont font parties les sphaignes que nous connaissons bien (Fig 1), ainsi que celui des Marchantiophyta, aussi appelées « hépatiques » que nous connaissons moins mais qui poussent parfois en compagnie de nos plantes carnivores en culture (Fig. 2).
Les Bryophytes ont une écologie bien différente des plantes à fleurs dont font partie toutes les plantes carnivores. Elles sont en effet extrêmement dépendantes de l’humidité directe du milieu pour leur croissance (étant donné qu’elles n’ont pas de racine) ainsi que pour leur reproduction car les gamètes mâles doivent nager dans une goutte d’eau pour rencontrer les gamètes femelles. Les Bryophytes sont souvent des espèces pionnières capables de pousser à même la roche ou le bois où les nutriments sont rares. Elles supportent généralement bien la sécheresse et peuvent se remettre en croissance rapidement une fois l’eau accessible.
Figure 2. Marchantia polymorpha est une hépatique à thalle que l’on retrouve parfois dans les pots des plantes carnivores. Les petits « palmiers » sont les structures reproductives sexuelles alors que les structures arrondies sont des sacs à propagules permettant la dissémination et la reproduction asexuée des individus. Photo : https://api.tela-botanica.org/img:002326044O.jpg
Des hépatiques… carnivores ?
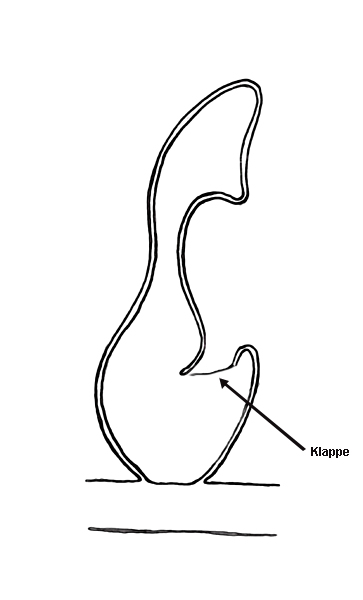
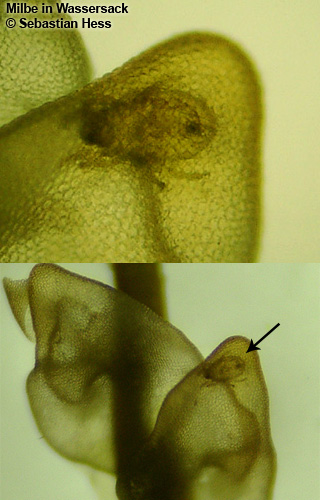
Vous avez bien lu, des hépatiques pourraient-elles être carnivores ? Deux espèces d’hépatique, Pleurozia purpurea (Fig. 3) et Colura zoophaga semblent être en mesure de capturer de petits protozoaires d’après deux études publiées en 2000 (Barthlott et al.) et 2005 (Hess et al.). Des chercheurs ont en effet mis en évidence la présence de « sacs », initialement destinés à stocker l’eau, dotés d’une petite ouverture qui ne s’ouvre que vers l’intérieur, ne permettant donc qu’une entrée et aucune sortie (Fig. 4). Ce « piège » ressemble en pratique à celui des Utricularia (mais sans mouvement) ou des Genlisea. Il semblerait aussi qu’il ne soit pas rare dans leur milieu naturel de trouver de petits protozoaires capturés à l’intérieur de ces pièges (Fig. 5). Des expérimentations en conditions contrôlées ont permis de montrer que des protozoaires étaient effectivement attirés par la plante sans que l’on arrive vraiment à comprendre comment ni pourquoi. En effet, 86% des vésicules contenaient des proies et certaines en contenaient plus de 10 ! La présence d’enzymes digestives est jugée hautement improbable par les auteurs mais n’a pas été clairement investiguée et statuée à l’heure actuelle.
Figure 3. Pleurozia purpurea, une des deux hépatiques à feuilles suspectées de carnivorie. Photo Dr. S. Hess.
Les expérimentations s’arrêtent malheureusement ici et on ne peut donc qu’émettre des hypothèses avec ces observations. La présence de ces protozoaires en milieu naturel est normale puisqu’ils se nourrissent des bactéries dégradant les parties mortes de la plante. Ils sont d’ailleurs plus nombreux sur les parties vieillissantes des mousses que sur les parties jeunes, ce qui corrobore cette hypothèse. Le mécanisme d’attraction de ces protozoaires autour des hépatiques est en revanche plus floue. Les auteurs précisent que cela peut venir du fait qu’une quantité importante de bactéries se trouve au niveau des pièges (sans que l’on ne sache pourquoi), ou bien que les protozoaires s’amoncellent autour de la plante pour se protéger de la lumière. Ces hypothèses alternatives n’ont malheureusement pas été testées. Une fois capturés, les protozoaires meurent dans les pièges mais ne semblent pas être digérés. Enfin, aucune expérience n’a été faite pour mesurer si la croissance des hépatiques était accélérée en présence de proies ou non, ce qui pose problème pour trancher la question de la carnivorie chez ces espèces. Comment souvent, nous avons plus de questions que de réponses. Mais finalement, quels sont les paramètres permettant d’affirmer qu’une plante est belle et bien « carnivore » ?
Figure 4. Schéma du piège de Pleurozia purpurea. Image : Dr. S. Hess.Figure 5. Protozoaire piégé par Pleurozia purpurea.
Joyeux bazar dans la carnivorie
Nous le savons bien, une plante est généralement considérée comme carnivore si elle parvient à attirer et capturer une proie, puis à la digérer afin d’en tirer des bénéfices sur sa croissance ou sa floraison, lui conférant ainsi une compétitivité et une reproduction accrues face aux autres espèces de son milieu. Le problème étant que bon nombre d’espèces considérées comme « carnivores » par les collectionneurs ne cochent pas toutes les cases. Par exemple, les Utricularia, et les Pinguicula dans une moindre mesure, ne semblent pas explicitement attirer des proies, mais comptent plutôt sur le hasard pour les piéger puis les digérer. De même, plusieurs espèces ne produisent pas d’enzymes digestives (comme certains Heliamphora) mais bénéficient tout de même de la capture de proies via des intéractions plus ou moins complexes avec d’autres espèces, notamment des bactéries. L’exemple le plus étudié étant Sarracenia purpurea qui contient tout un écosystème à l’intérieur de ses pièges permettant la dégradation des proies (Harvey & Miller, 1996).
On pourrait alors penser que le seul point commun entre toutes les plantes carnivores serait la production de feuilles extrêmement modifiées dans l’unique but de capturer des proies. Cela serait logique d’un point de vue évolutif, si la sélection naturelle a favorisé le développement de feuilles qui se détournent de leur utilité première (optimiser la captation de la lumière solaire pour la photosynthèse) au profit de la création de formes et de mécanismes complexes pour la capture d’animaux, alors nécessairement l’espèce a un penchant pour la carnivorie. Mais alors, que faire de Paepalanthus bromélioides qui est capable d’absorber les nutriments des animaux qui se noient au centre de sa rosette (Nishi et al, 2013), ou de Brocchinia reducta qui a les mêmes capacités via des glandes spécialisées (Owen et al, 1988) et qui semble en plus produire des enzymes digestives (phosphatases) en faible quantité (Płachno et al., 2006). En effet, ces deux espèces généralement considérées comme carnivores ou protocarnivores sont en tous points similaires à leurs congénères pour lesquels ces attributs n’existent pas (Fig. 6.).
La modification drastique des feuilles n’est peut-être pas nécessaire à la carnivorie, le plus important étant peut-être la finalité ultime du syndrome carnivore : récupérer les nutriments d’autres organismes pour améliorer sa compétitivité dans un milieu pauvre. Si l’on y réfléchit bien, c’est tout de même là l’intérêt de la carnivorie. Donc, si une espèce est capable d’absorber des nutriments et de les intégrer à ses tissus, elle pourrait être qualifiée de « carnivore ». Ça tombe bien, une expérience permet justement de marquer des protéines déposées à la surface des feuilles d’une plante et d’en mesurer la quantité qui intègre les tissus du végétal. Cette expérience a mené à quelques surprises. Par exemple, chez les Stylidium, les nutriments marqués ne sont pas plus absorbés par les espèces de ce genre que des espèces aléatoires – non carnivores – et le niveau d’absorption serait d’ailleurs nettement inférieur à celui des Drosera (Nge & Lambers, 2018). Autre surprise, plusieurs espèces qui sécrètent du mucilage (à la manière des plants de tomates par exemple), mais usuellement considérées comme « non carnivores » par les collectionneurs, absorbent en réalité les nutriments des proies qu’elles capturent (Spomer, 1999), faisant d’elles des plantes « davantage » carnivores que les Stylidium. En effet, d’après Spomer (1999), 13 espèces collantes sur les 17 testées sont capables de digérer des nutriments via des protéases, sans que l’on ne sache si cela apporte réellement un avantage à la plante. Dans son article, Spoomer montre que la patate (Solanum tuberosum) est carnivore, et que ses feuilles digèrent les protéines. L’intégration des protéines aux tissus de la plante n’a été testé que chez Geranium viscosissimum et Potentilla arguta et a été largement validé (Fig. 7). Ce sont donc aussi des plantes carnivores.
La digestion et l’absorption de nutriments issus de proies piégés pourraient alors être bien plus répandues qu’on ne l’imaginait chez les plantes collantes. Il est pourtant largement admis que le mucilage sert avant tout de protection faces aux phytophages. Ces espèces pourraient alors faire d’une pierre deux coups en se protégeant des ravageurs tout en absorbant les nutriments de ceux qui se font piéger, démontrant ainsi une piste plausible de l’évolution des mécanismes de capture des pièges à glu. Mais une question demeure, si la plante ne tire pas de bénéfices notables sur sa compétitivité, peut-on vraiment parler de carnivorie ? En effet, ces espèces ne semblent pas pousser sur des milieux spécialement pauvres (Chase et al., 2009).
Figure 7. Geranium viscosissimum. Cette espèce absorbe les nutriments via les proies qu’elle capture au niveau de ses feuilles, ses tiges et de ses calices glanduleux. En revanche, il n’a pas été testé si la plante gagnait un avantage compétitif sur ses congénères si elle était « nourrie ». Photo : https://en.wikipedia.org/wiki/Geranium_viscosissimum#/media/File:Geraniumviscosissimum.jpg
Il est clairement plus compliqué que prévu de déterminer si une plante peut être considérée comme « carnivore » ou non, et nous ne nous attarderons même pas sur le terme de « protocarnivore » qui englobe un peu tout et n’importe quoi sans consensus claire. Un autre exemple pour continuer à vous casser le cerveau si vous aviez encore des certitudes à propos de la définition de la carnivorie végétale. L’espèce Capsella bursa-pastoris (ou la bourse-à-Pasteur, une jolie Brassicaceae qui pousse probablement près de chez vous et qui produit des siliques en forme de cœur) produit des graines qui semblent pouvoir créer du mucilage une fois au contact de l’eau afin de piéger de petits nématodes (Fig. 8), permettant ainsi aux graines de mieux germer et d’accumuler plus de réserves, et aux plantules de développer des feuilles plus grandes (Roberts et al., 2018). Cette capacité semble facultative puisqu’elle n’est observée que quand les sols sont pauvres en nutriments. La plante en soit n’est donc pas carnivore, sa graine en revanche semble pouvoir l’être mais uniquement dans certaines conditions. Peut-on alors considérer l’espèce comme carnivore ?
Figure 8. Graines de Capsella bursa-pastoris attirant des nématodes. Photo Roberts et al., 2018.
On commence là à s’éloigner de la dionée qui semble calibrée pour capturer et digérer des proies, pourtant il ne semble pas y avoir de différences fondamentales sur le principe entre la graine de la bourse-à-Pasteur et la dionée. Néanmoins, toutes les plantes ne sont pas carnivores, malgré le fait qu’elles puissent « capturer » ou plutôt tuer des animaux, quoiqu’en disent les informations qui circulent sur les réseaux sociaux. Prenons l’exemple de Puya raimondii, la reine des Andes, que j’ai eu la chance d’observer dans son milieu naturel dans les hauts-plateaux boliviens (Fig. 9). Cette espèce majestueuse a souvent été suspectée de carnivorie (ou de protocarnivorie) à cause d’intéractions qu’elle aurait avec des oiseaux, ou le fait qu’elle puisse tuer des animaux (Rees & Roe, 1980). Des photos montrent même des chats sauvages morts entortillés dans ses feuilles (je vous laisse chercher le lien dans les références). Cette hypothèse stipule que les déjections des oiseaux et les animaux morts piégés dans ses feuilles épineuses enrichissent le sol une fois dégradés, permettant ainsi une meilleure croissance de la plante.
Pourquoi cette hypothèse, néanmoins séduisante, reste bancale ? Tout d’abord il faut savoir que cette espèce pousse sur les hauts plateaux andins à plus de 4’000 mètres d’altitude, dans un climat particulier où les gelées sont courantes la nuit et le soleil est brûlant durant la journée. Les précipitations sont rares et le sol plutôt pauvre ce qui rend difficile la dégradation de la matière organique (on observe le même phénomène dans les sols de nos montagnes). Difficile donc a priori d’imaginer une transformation efficace des animaux piégés par la plante en nutriments absorbables même au terme de sa longue vie. La végétation dominante appelée « pampa » est composée quasiment exclusivement de graminées dépassant rarement les quelques dizaines de centimètre de hauteur. La plante suspectée de carnivorie peut en revanche atteindre 15 mètres de haut en floraison et représente souvent l’unique point en hauteur du paysage. Elle pourrait alors faire office de perchoir, expliquant ainsi la présence des oiseaux. De plus, les épines et la dureté de ses feuilles sont des syndromes récurrents de protection face aux herbivores, surtout dans un milieu où l’eau est rare et les graminées peu appétissantes. Le fait que des animaux se retrouvent piégés dans ses feuilles montre en réalité à quel point cette défense est efficace. Cela explique aussi pourquoi les oiseaux viennent nidifier à l’intérieur de celles-ci puisqu’elles offrent une protection diablement efficace face aux prédateurs. D’ailleurs, on peut penser que les chats retrouvés morts venaient chasser les oiseaux réfugiés à l’intérieur de la plante, qui leur a sauvé la vie. Dans cette situation, on ne peut donc pas parler de carnivorie mais simplement de mécanismes de défense face aux herbivores.
Figure 9. Puya raimondii observé dans l’altiplano bolivien en 2015, moi-même pour l’échelle.
Trier les syndromes carnivores et la carnivorie
Il est toujours très compliqué de ranger les éléments naturels dans des cases bien définies et le cas de la carnivorie ne fait pas exception. Nous n’avons même pas parlé ici des espèces pouvant dégrader des végétaux (Nepenthes ampullaria), qui s’apparenteraient en toutes logiques à des plantes « herbivores », ou des espèces ayant des intéractions bien spécifiques avec des animaux pour tirer des nutriments de leurs déjections, que l’on pourrait appeler plantes « détritivores », pour rester poli (on en avait déjà parlé dans cet article). Il est très important de se rappeler que, comme toujours, il y a des intermédiaires, des exceptions et que tout est une question de nuances et de définition. Une plante est-elle « carnivore » si seulement sa graine arrive à tirer des bénéfices de la capture d’animaux, uniquement dans des conditions bien particulières ? Une plante est-elle « carnivore » si elle arrive à absorber les nutriments de ses proies sans pour autant en tirer un bénéfice notable sur sa croissance ? Une plante est-elle « protocarnivore » si elle ne digère pas elle-même les proies qu’elles capturent mais qu’elle tire des bénéfices notables sur sa croissance grâce à l’intervention de bactéries ou d’animaux ?
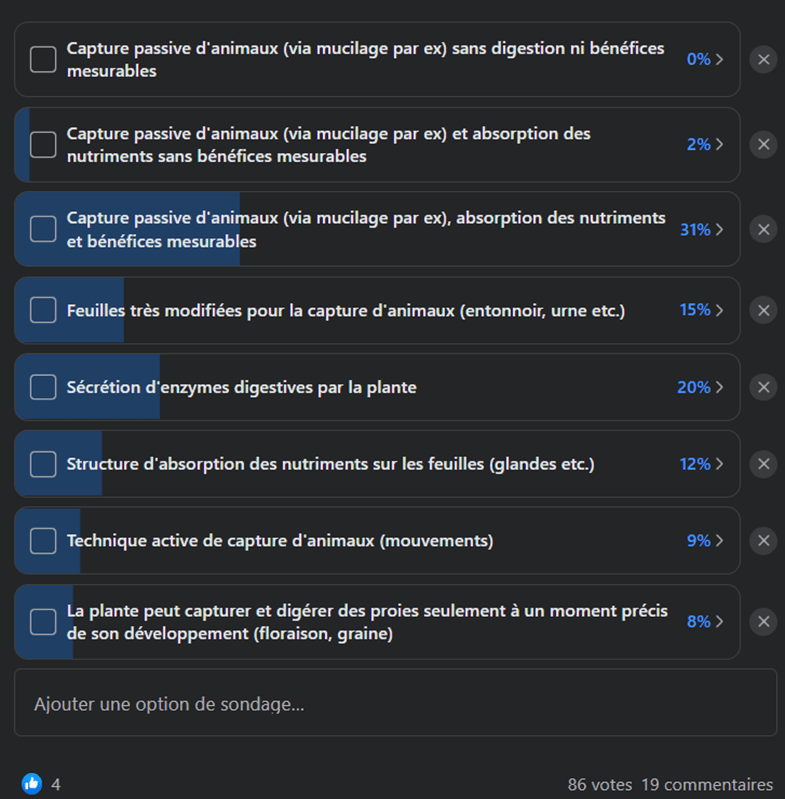
C’est compliqué, et à chacun de se faire son avis. Et en parlant d’avis, voici le résultat d’un sondage qui cherchait à déterminer, selon les utilisateurs d’un groupe Facebook, quelles propositions pouvaient définir une plante carnivore (Fig. 10). Les utilisateurs pouvaient voter pour plusieurs réponses simultanément car elles ne sont pas exclusives pour la plupart. Pourtant on voit que la réponse la plus sélectionnée peine tout de même à réunir un tier des 86 votants, ce qui montre qu’il n’y a pas vraiment de consensus, même au sein de la communauté (sinon elle aurait été proche de 100%). Les critères qui semblent particulièrement importants reposent sur les bénéfices tirés de l’absorption des nutriments des proies et la sécrétion d’enzymes digestives, les autres réponses ne convenant pas à plus d’une personne sur cinq.
Figure 10. Résultats du sondage effectué sur un groupe Facebook en Septembre 2022 regroupant 86 votants.
La sélection au cours de l’évolution de l’habilité à sécréter des enzymes digestives semble être un réel syndrome carnivore, sinon il est difficile de l’expliquer. De même, la finalité de la carnivorie est de tirer des bénéfices sur la compétitivité des individus en absorbant plus de nutriments que ses voisins. Pourtant, plusieurs espèces ne produisent pas d’enzymes digestives mais bénéficient pour autant de la capture de leurs proies, alors comment définir clairement la carnivorie ?
Pour tenter de terminer sur une note plus constructive que « c’est compliqué », quelques auteurs ont tenté des définitions avec à chaque fois des points forts et des points faibles. Certains séparent la « vraie » carnivorie intégrant l’absorption de nutriments issus de proies, de la « zoophagie » qui ne s’applique qu’aux plantes capables de capturer des animaux peu importe la raison profonde (ce qui s’apparenterait à un caractère « protocarnivore »), sans considérer le gain de compétitivité. Certains auteurs parlent aussi de carnivorie « active » et « passive » en fonction de l’intensité des mécanismes mis à disposition par la plante pour tirer des bénéfices de ses proies sans tracer une délimitation claire entre les deux. D’autres évoquent le terme de « plantes meurtrières » pour les regrouper toutes sous un même terme sans chercher de définition précise à la carnivorie (Chase et al., 2009). Enfin, d’autres partent du principe que la sécrétion d’enzymes digestives est l’unique critère permettant de définir si une plante est carnivore ou non, ce qui a le mérite d’être clair, mais reste discutable. En effet, Sarracenia purpurea n’est donc pas carnivore selon cette définition alors même que ses feuilles sont extrêmement modifiées pour capturer des proies et absorber leurs nutriments. On peut alors se demander si la modification des feuilles est réellement un attribut carnivore contrairement à la sécrétion d’enzymes digestives et on retombe sur les premières réflexions… C’est un peu le serpent qui se mord la queue.
D’un point de vue purement personnel, et ce qui suit n’engage que moi, je trouve intéressant de considérer la carnivorie non pas comme un critère binaire (« carnivore » ou « non carnivore ») mais plutôt comme une échelle, une nuance ou une graduation, en fonction des mécanismes mis en place par la plante et de sa dépendance aux proies. Il n’y aurait donc pas de limite claire entre une plante « carnivore » et une qui ne le serait pas, juste des niveaux de carnivorie (ou de « syndrome carnivore ») plus ou moins élevés qui reflètent en quelque sorte l’intensité de la carnivorie d’un point de vu évolutif. Voici une ébauche :
0 = Pas de syndrome carnivore.
1 = Mécanismes de défense face aux herbivores tel que le mucilage, ou mécanismes entraînant la mort d’animaux sans absorption des nutriments (Plumbago capensis par exemple, Fig. 11). On peut considérer les plantes de ce niveau comme « zoophages ».
2a = Absorption claire de nutriments issus de la capture de proies sans mécanisme dédié et sans avantage clair sur les autres espèces du milieu. A priori, je n’ai pas trouvé de plantes capables d’absorber des nutriments sans mécanismes dédiés (glandes, poils etc.), cela peut être le cas chez Geranium viscosissimum ou Paepalanthus bromelioides mais ça n’a pas été clairement investigué.
2b = Absorption claire de nutriments issus de proies via des glandes spécialisées ou autre mécanisme dédié sans avantage clair sur les autres espèces du milieu (Brocchinia reducta peut faire partie de ce niveau de carnivorie ou du suivant, je ne crois pas que l’amélioration de la compétitivité ait été démontrée). On pourrait parler de protocarnivorie pour l’ensemble du niveau 2.
3 = Avantage compétitif et/ou reproductif démontré à l’absorption de nutriments issus de proies (Capsella bursa-pastoris et ses graines carnivores). Les espèces de ce niveau seraient alors « faiblement carnivores ».
4 = Organes nettement modifiés pour l’attraction et/ou la capture de proies indiquant une évolution franche vers la carnivorie ; absorption des nutriments entrainant une augmentation de la compétitivité (Sarracenia, Nepenthes, etc.). Ce niveau et le suivant regroupent ce que l’on appelle communément les plantes « carnivores ».
5 = Mêmes syndromes que le point 4 mais avec en plus des modifications physiologiques complexes facilitant la carnivorie, par exemple la présence de mouvements favorisant la capture ou la digestion (Drosera, Utricularia, Dionaea etc.). Les espèces de ce niveau seraient alors « très carnivores ».
Figure 11. Glandes sécrétant du mucilage au niveau des sépales de Plumbago capensis (= Plumbago auriculata). Une sécrétion de protéase a pu être observée mais aucune absorption de nutriments n’a été démontré (Stoltzfus et al., 2002).
Concluons
Toutes ces réflexions sur la carnivorie sont très théoriques et aucune n’est tout à fait satisfaisante pour l’ensemble des végétaux concernés. Dans l’échelle proposée, l’hypothèse de départ stipule que la sécrétion d’enzymes digestives ou la digestion des proies via des agents externes (bactéries) sont des mécanismes similaires puisqu’ils amènent à une même finalité qui est l’absorption de nutriments. Cette hypothèse est discutable mais néanmoins logique dans le sens où une espèce n’a pas « besoin » de développer tout un métabolisme de sécrétion d’enzymes digestives si le travail de digestion est déjà fait par d’autres organismes. De plus, des espèces de niveau 1 ou 2 dans l’échelle proposée semble en mesure de sécréter des protéases, mais il n’est pas très clair si ces enzymes proviennent bel et bien de la plante ou de bactéries à la surface de ses organes.
Pour en revenir à nos hépatiques (vous vous souvenez ?), il est difficile de les qualifier de « carnivores » à l’heure actuelle vu que nous ne savons pas si elles absorbent les nutriments de leurs proies et si elles en tirent un avantage compétitif. Elles sont à minima « zoophage » (au niveau 1 de l’échelle) mais peuvent tout à fait se retrouver au niveau 4 s’il est démontré qu’elles peuvent absorber les nutriments induisant des bénéfices nets sur leur compétitivité, étant donné qu’elles possèdent des organes qui semblent dédiés à la capture d’animaux.
Pour conclure, il est tout de même intéressant de noter que si des espèces très ancestrales ou « primitives » comme des mousses (ne dîtes jamais ça à un/e bryologue) ont des capacités d’attraction et de capture d’animaux, alors la carnivorie chez les plantes a pu potentiellement évoluer depuis le tout début de la colonisation des terres par les végétaux, ce qui expliquerait qu’elle soit apparu au moins une dizaine de fois au cour de l’évolution de manière tout à fait indépendante rien que chez les plantes à fleurs, qui sont relativement récentes. Pour aller plus loin dans ces réflexions, il nous manque avant tout des recherches dans le domaine pour mieux « classifier » la carnivorie en fonction des attributs des plantes. Il faudrait plus largement investiguer la présence de structures spécialisées dans l’absorption de nutriments, la capacité des plantes à intégrer les nutriments des proies dans leurs tissus et les potentiels bénéfices sur leur croissance et reproduction pour avoir un avis plus éclairé sur bon nombre d’entre-elles. D’un point de vue évolutif, il serait intéressant d’étudier si la sécrétion d’enzymes digestives est un attribut antérieur à la modification des feuilles, ou au contraire un caractère plus « compliqué » à mettre en place et donc réservé à des plantes franchement carnivores. Quoi qu’il en soit, le petit monde des plantes carnivores semble avoir des limites floues et pourrait tout à fait inclure tout un tas de nouvelles espèces, au plus grand bonheur des collectionneurs.
Pour aller plus loin
Un article scientifique qui traite de la carnivorie d’une bonne partie des plantes citées dans cette article.
Chase, M. W., Christenhusz, M. J., Sanders, D., & Fay, M. F. (2009). Murderous plants: Victorian Gothic, Darwin and modern insights into vegetable carnivory. Botanical Journal of the Linnean Society, 161(4), 329-356. https://academic.oup.com/botlinnean/article/161/4/329/2418471
Harvey, E., & Miller, T. E. (1996). Variance in composition of inquiline communities in leaves of Sarracenia purpurea L. on multiple spatial scales.Oecologia, 108(3), 562-566.
Nge, F. J., & Lambers, H. (2018). Reassessing protocarnivory–how hungry are triggerplants?. Australian Journal of Botany, 66(4), 325-330. https://www.publish.csiro.au/BT/BT18059
Nishi, A. H., Vasconcellos-Neto, J., & Romero, G. Q. (2013). The role of multiple partners in a digestive mutualism with a protocarnivorous plant. Annals of Botany, 111(1), 143-150. https://academic.oup.com/aob/article/111/1/143/182710
Owen Jr, T. P., Benzing, D. H., & Thomson, W. W. (1988). Apoplastic and ultrastructural characterizations of the trichomes from the carnivorous bromeliad Brocchinia reducta. Canadian journal of botany, 66(5), 941-948. https://cdnsciencepub.com/doi/abs/10.1139/b88-136
Rees, W. E., & Roe, N. A. (1980). Puya raimondii (Pitcairnioideae, Bromeliaceae) and birds: an hypothesis on nutrient relationships. Canadian Journal of Botany, 58(11), 1262-1268. https://cdnsciencepub.com/doi/abs/10.1139/b80-157
Roberts, H. R., Warren, J. M., & Provan, J. (2018). Evidence for facultative protocarnivory in Capsella bursa-pastoris seeds. Scientific Reports, 8(1), 1-5. https://www.nature.com/articles/s41598-018-28564-x



















{kind=link}
{kind=link}
{kind=link}